



Функция щитовидной, половых желез, коры надпочечников регулируется гормонами передней доли гипофиза (аденогипофиза).
Общее название этих гормонов – тропные гормоны:
С некоторой условностью к тронным гормонам относится и соматотропный гормон гипофиза (гормон роста), который оказывает влияние на рост непосредственно, но и опосредовано через гормон соматомедин, образующийся в печени.
Кроме того, в аденогипофизе образуется интермедии (меланоцитстимулирующий гормон – МСГ) и пролактин которые обладают прямым эффектом на периферические органы.
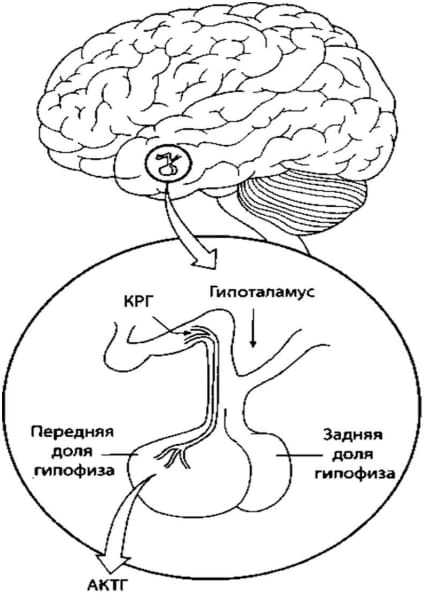
Рисунок 1. Гипоталамо-гипофизарная система
В свою очередь, высвобождение всех 7 гормонов аденогипофиза зависит от гормональной активности нейронов гипофизотропной зоны гипоталамуса – ее медиальной области (в основном паравентрикулярное ядро).
Здесь образуются гормоны, оказывающие стимулирующее и тормозящее влияние на секрецию гормонов аденогипофиза. Стимуляторы называются рилизинг-гормонами (либеринами), ингибиторы – статинами.
Гипоталамус получает информацию о состоянии внутренней среды по нескольким каналам:
+ афферентные возбуждения поступают в мозг от экстеро- и интерорецепторов;
+ несинаптическая диффузная афферентация реализуется путем дистантного (через кровь) действия медиаторов и других биологически активных веществ.
По функциональному принципу нейрогормоны гипоталамуса разделяют на 3 группы:
Висцеротропные гормоны – вазопресснн и окситоцин – оказывают прямое действие на некоторые периферические железы внутренней секреции и в основном на неэндокринные органы, осуществляющие вегетативные функции: почки, сердце, сосуды, репродуктивные органы, включая молочные железы.
Аденогипозотропные гормоны либерины (рилизинг-гормоны) и статины регулируют гормональную функцию аденогипофиза и через него секреторную активность различных желез внутренней секреции.
Рилизинг-гормоны освобождаются из отростков нервных клеток, тела которых лежат во многих ядрах гипоталамуса, наибольшая плотность их отмечается в паравентрикулярном ядре.
Регуляция гормональной активности большинства желез внутренней секреции осуществляется но принципу отрицательной обратной связи: сам гормон, его количество в крови, регулирует свое образование. Такое воздействие опосредуется через образование соответствующих рилизинг-гормонов.
Например, при повышении в крови уровня гормона коры надпочечников – кортизола в срединном возвышении высвобождается меньше АКТГ-РГ, в результате чего секреция АКТГ гипофизом снижается; что приводит к уменьшению образования кортикостероидов надпочечниками и снижению содержания кортизола в крови.
Нейроны гипоталамуса, образующие рилизинг-гормоны (РГ), являются своеобразными клетками с двойной функцией: с одной стороны, это типичный нейрон, интегрирующий на себе влияние других отделов ЦНС и гуморальных регуляторов, а с другой – эндокринная клетка.
Гипоталамус имеет характерную отличительную особенность – это слабо выраженный гематоэнцефалический барьер.
Благодаря этому из протекающей крови выходят гормоны и другие биологически активные соединения, которые и оказывают – свое регулирующее влияние на нейроны.
Данный механизм саморегуляции сохраняется в отсутствие влияний на гипоталамус со стороны ЦНС (после полного отделения медиальной области гипоталамуса от остальных отделов ЦНС).
Но в естественных условиях другие отделы ЦНС участвуют в приспособлении этой реакции к внутренним и внешним потребностям организма. Например, образование кортизола корой надпочечников резко увеличивается при стресс-реакциях, этому предшествует увеличение секретна AKTГ-PГ и выхода АКТГ.
Сигналы от этих центров к ядрам гипоталамуса передаются посредством нейронов, относящихся к моноаминергическим путям головного мозга, медиаторами которых являются биогенные амины – норадреналин, серотонин, дофамин.
К указанным центрам ЦНС поступают не только нервные импульсы, но и информция об уровне гормонов в крови, а они через моноаминовые пути оказывают соответствующее влияние на специфические структуры гипоталамусы и на выработку или РГ, то есть модулируют образование РГ, поскольку на мембране нейронов указанных отделов кроме обычных синапсов имеются также рецепторы к различным гормонам.
Выведение и распространение гипоталамических нейрогормонов осуществляется 3 путями.
Трансадентгипофизарный путь: гипоталамус контролирует аденогипофиз, в первую очередь выработку его тронных гормонов.
Таким образом он регулирует функции периферических гипофиззафисимых желез внутренней секреции, а также рост и дифференцировку различных тканей, секреторную функцию экзокринных гормонозаивсимых желез (молочной), образование пигмента.
Из мозговой жидкости нейрогормоны поступают также в ткань мозга и в сосуды воротной системы, несущие кровь к аденогипофизу.
Нейрогормоны, поступающие из мозговой жидкости в ткань мозга, аденогипофиз и общий кровоток, осуществляют нейротропное, гипофизотропное и висцеротропное действие.
Параденогипофизарный путь прямой гипоталамической регуляции, минуя аденогипофиз, включает:
= дистантную регуляцию внутренних органов (сосуды, матка, эпителий почечных канальцев, молочные железы). Вазопрессин и окситоцин из гипоталамуса поступают в нейрогипофиз и далее в общий кровоток; достигают клеток-мишеней, где стимулируют клеточные реакции;
= поступление высоких концентраций этих гормонов в кровь из депо нейрогипофиза при стрессе и некоторых заболеваниях. В высоких концентрациях в условиях стресса они выполняют защитную функцию, так как ограничивают избыточную продукцию гормонов, в частности кортикостероидов, надпочечниками и предупреждают «истощение» желез, внутренней секреции.
Гипофиз расположен в гипофизарной ямке турецкого седла основной кости. При помощи ножки он связан с основанием мозга, состоит из передней доли (аденогипофиза) и задней доли – нейрогипофиза.
В передней доле гипофиза вырабатываются тропные и эффекторные гормоны. Тропные гормоны: тиреотропный гормон (тиреотропин), адренокортикотропный гормон (кортикотропин), гонадотропные гормоны (гонадотропины); они регулируют деятельность периферических желез внутренней секреции.
Аденогипофиз является комплексом 6 желез, каждая из которых состоит из особых клеток, контролируемых по-своему. Все клетки получают стимулирующие сигналы лишь гуморальным путем, реагируя на пептиды гипоталамуса и катехоламины (дофамин, норадреналин, гистамин).
В процессах секреции гормонов принимают непосредственное участие вторые посредники, особенно ионы кальция и цАМФ.
Тропные гормоны передней доли гипофиза:
* Тиреотропный гормон (тиреотропин) стимулирует функцию щитовидной железы, вызывает гиперплазию ее железистой ткани, стимулирует выработку тироксина и трийодтиронина.
* Адренокортикотропный гормон (кортикотропин) стимулирует функцию пусковой зоны корм надпочечников, в которой образуются гормоны глюкокортикоиды и приводит к увеличению образования этих гормонов.
Продукция АКТГ подвержена суточной периодичности, что связано с ритмичностью выделения кортиколиберина. Максимальные концентрации АКТГ отмечаются утром в 6-8 часов, минимальные – с 18 до 23 часов.
Образование АКТГ регулируется кортиколиберином гипоталамуса. Секреция АКТГ усиливается при стрессе и факторах, вызывающих стрессогенные состояния: холод, боль, физические нагрузки, эмоции.
* Гонадотропные гормоны (гонадотропины).
Фолликулостимулирующий гормон (фоллитропин, ФСГ) стимулирует рост и созревание везикулярного фолликула в яичнике и их подготовку к овуляции. Влияние фоллитропина на образование женских половых гормонов (эстрогенов) небольшое.
Этот гормон имеется как у женщин, так и у мужчин. У мужчин под влиянием фоллитропина происходит образование половых клеток (сперматозоидов).
Лютеинизирующий гормон (лютропин, ЛГ) необходим для роста везикулярного фолликула яичника на стадиях, предшествующих овуляции, и для самой овуляции (разрыв оболочки созревшего фолликула и выход из него яйцеклетки), образования желтого тела на месте лопнувшего фолликула.
Лютропин стимулирует образование женских половых гормонов (эстрогенов). Однако, для того, чтобы этот гормон осуществил свое действие на яичник, необходимо предварительное длительное действие фоллитропина.
Секреция ФСГ и ЛГ регулируется гонадолиберином гипоталамусса. Образование гонадолиберина, ФСГ и ЛГ зависит от уровня эстрогенов и андрогенов и регулируется по механизму обратной связи.
Гормон аденогипофиза пролактин угнетает продукцию гонадотропных гормонов. Тормозное действие на выделение Л Г оказывают глюкокортикоиды.
Эффекторные гормоны передней доли гипофиза.
Наиболее ярко влияние гормона выражено на костную и хрящевую ткани. Под влиянием роста эпифизарных хрящей в длинных костях верхней и нижней конечностей происходит рост костей в длину.
Гормон роста усиливает секрецию инсулина посредством соматомединов, которые образуются в печени; влияет на углеводный обмен, оказывая инсулиноподобное действие; усиливает мобилизацию жира из депо и использование его в энергетическом обмене.
Гормон стимулирует синтез белка – лактальбумина, жиров и углеводов молока. Пролактин обладает также и лютеотропным действием (способствует продолжительному функционированию желтого тела и образованию им гормона прогестерона).
Пролактин влияет на водно-солевой обмен организма, задерживая воду и натрий в организме; усиливает эффекты альдостерона и вазопрессина, повышает образование жира из углеводов. Образование пролактина регулируется пролактолиберином и пролактостатином гипоталамуса.
Гормоны задней доли гипофиза. Эти гормоны образуются в гипоталамусе, в нейрогипофизе происходит их накопление. В клетках супраоптического и паравентрикулярного ядер гипоталамуса осуществляется синтез окситоцина и антидиуретического гормона.
В нейронах паравентрикулярного ядра образуется преимущественно гормон окситоцин, а в нейронах супраоптического ядра – вазопрессин (антидиуретический гормон).
Синтезированные гормоны путем аксонального транспорта с помощью белка переносчика нейрофизита по гипоталамо-гипофизарному тракту транспортируются в заднюю долю гипофиза. Здесь происходит депонирование гормонов и дальнейшее выделение в кровь.
Антидиуретический гормон (АДТ) или вазопрессин осуществляет в организме 2 основные функции:
# Антидиуретическое действие заключается в стимуляции реабсорбции воды в дистальном отделе нефрона и осуществляется благодаря взаимодействию гормона с вазопрессиновыми рецепторами, что приводит к повышению проницаемости стенки канальцев и собирательных трубочек для воды, ее реабсорбции и концентрированию мочи.
В клетках канальцев происходит также активация гиалуронидазы, что приводит к усилению деполимеризации гиалуроновой кислоты, в результате чего повышается реабсорбция воды и увеличивается объем циркулирующей жидкости.
# В больших (фармакологических) дозах АДГ суживает артериолы, в результате чего повышается артериальное давление, поэтому его также называют вазопрессином. В обычных условиях при его физиологических концентрациях в крови это действие не имеет существенного значения.
Однако, при кровопотере, болевом шоке происходит увеличение выброса АДГ. Сужение сосудов в этих случаях может иметь адаптивное значение.
Окситоцин избирательно действует на гладкую мускулатуру матки, вызывая ее сокращение при родах. На поверхностной мембране клеток существуют специальные окситоциновые рецепторы.
Во время беременности окситоцин не повышает сократительную активность матки, но перед родами под влиянием высоких концентраций эстрогенов резко повышается чувствительность матки к окситоцину.
Увеличение секреции окситоцина происходит под влиянием импульсов от рецепторов шейки матки, а также механорецепторов сосков грудной железы при кормлении грудью. Эстрогены усиливают секрецию окситоцина.
Функции окситоцина в мужском организме изучены недостаточно, считают, что он является антагонистом АДГ.
(Кузенко, С.С. Тревожные расстройства: диагностика, психофармакология и психотерапия: учебное пособие, Экоинвест)






