Тропические биомы
Положение. Тропические биомы занимают 22% поверхности Мирового океана, их границы примерно совпадают с 30° параллелью в каждом океане.
Поверхностный слой имеет температуру 28–29°С. Годовая разница между наибольшей и наименьшей температурой не превышает один градус. Подповерхностный слой существенно холоднее. В восточных частях океанов на глубине 35–40 м температура падает до 16°С. Лишь в центральных частях субтропических круговоротов, где происходит опускание воды, стратификация выражена плохо.
Таксономическое разнообразие тропических биомов наиболее велико. Ареалы тропических видов обычно циркумтропические, при более узких ареалах, они обычно охватывают или Индийский и Тихий океаны, или Атлантический.
Фитопланктон. В тропических биомах существуют два типа годовой динамики первичной продукции: постоянно низкая и зависящая от муссонов (см. выше). Первичная продукция низка из-за постоянной стратификации, ведущей к постоянному же дефициту биогенов. Поэтому основные продуценты — мелкие водоросли (пико-планктон), дающие до 90% биомассы и до 80% первичной продукции.Уровень первичной продукции в течение года на большей части площади тропических биомов меняется незначительно. Лишь в районах, подверженных действию муссонов есть аналог биологических сезонов Sverdrup’а. В начале каждого из двух периодов муссонов происходит смена направления ветра на противоположный (это и называют муссоном).
Происходит также заглубление пикноклина, хотя и значительно меньшее, чем в биомах западных ветров, всё же пикноклин часто опускается ниже компенсационной глубины. Это приводит к появлению четырёх сезонов в динамике первичной продукции.
Наиболее отчётливо муссонная динамика выражена на севере Индийского океана. В тропическом биоме широко развита фиксация азота атмосферы. Основные азотфиксаторы — цианобактерии Trichodesmum spp., их распространение ограничено сезонной изотермой 20°С. В фиксации азота существенную роль играют некоторые диатомовые, имеющие эндосимбиотические цианобактерии.
Постоянно низкая первичная продукция способствует развитию разнообразных симбиотрофных связей, таких как пластиды водорослей, функционирующие в Heliozoa после того, как они съедят сами водоросли.
Зоопланктон. Стабильность вертикальной гидрологической структуры и равномерное распределение первичной продукции в течение года, позволяет формироваться видам, свойственным отдельным слоям: плейстону, поверхностному и подповерхностному.
В результате для низких широт характерно очень высокое таксономическое разнообразие всех групп. Вертикальные миграции мелких фитофагов отсутствуют, редко слаборазвиты суточные миграции. Из-за отсутствия миграций мелких фитофагов более выражены миграции следующего трофического уровня: макропланктона и нектона (кальмары, ракообразные, рыбы.Хотя вертикальные миграции зоопланктона существуют везде, в тропических биомах они особенно отчётливы: существенная часть макропланктона и нектона, особенно крупные копеподы, эуфаузиид, светящихся анчоусов Myctophidae и кальмаров поднимается к поверхности вечером и опускается вниз на 200–500 м (до 1500 м) днём.
Даже такой крупный представитель планктона, как луна-рыба, питается преимущественно на глубине 110–180 м, затем поднимается к поверхности (0–5 м), где согревается после охлаждения на глубинах. Ежедневные вертикальные миграции фитофагов позволяют фитопланктону за день восстановить свою численность, поэтому фактическая продуктивность в тропических биомах ближе к потенциальной, чем в прочих, где выедание более постоянно во времени. Мигрирующий макрозоопланктон и нектон создаёт звукорассеивающие слои.
Доля хищного зоопланктона от 20–25% в полярных районах возрастает до 40–55% в тропических биомах, около трети этого роста обеспечивают виды, питающиеся желетелыми. Ведущие таксоны зоопланктона — копеподы и эуфаузииды — дают от половины до 70% биомассы (оценка по содержанию углерода).
Рыб, питающихся фитопланктоном, нет, по-видимому, из-за очень низкой его концентрации и мелких размеров его клеток. Планктонных чисто эпипелагических рыб мало: лишь карликовые виды макрелещуковых и некоторых других. Гораздо больше планктонных рыб, совершающих вертикальные миграции. Это уже упоминавшиеся светящиеся анчоусы и близкие к ним семейства.
Нейстон и пелагическое обрастание. Как отмечено выше, настоящие плейстонные виды (обитающие в приповерхностном слое воды или на её поверхности постоянно (Рис. 6.18)) есть только в тропических биомах, хотя плейстонные стадии в развитии видов есть и в других биомах.
Другие — на ней (клопы-водомерки Halobates), их называют эпиплейстон. Третьи частично находятся над поверхностью воды, частично — под ней, как хондрофоры, сифонофора Physalia aretusa. В состав плейстона входят даже рыбы: Nomeus — комменсал кишечнополостных, обитающий ду их щупальцами, луна-рыба выставляет из воды спинной плавник и использует его как парус.
Структуру ассоциаций плейстона создают хондрофоры и Physalia. И те, и другие имеют симбиотические зооксантеллы. Они образуют скопления протяжённостью в сотни миль и плотностью до 1 экз./кв. м и даже более. Их щупальца опускаются на глубину до 10 м и более. Учитывая, что плейстон характерен для олиготрофных районов тропических круговоротов, фактически он — один из существенных продуцентов.
В состав нейстона входят и уже упоминавшиеся маты диатомовых водорослей.
Генетическая связь между ними и донными сообществами отсутствует. Обрастание это формируется на длительное время плавающих по поверхности брёвнах и других аналогичных с точки зрения биоты предметах (кусках пемзы, а ныне — буях и судах).
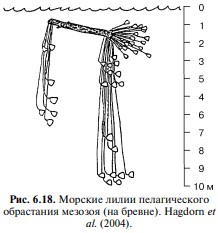
Несмотря на кажущуюся эфемерность и экзотичность брёвен, судя по тому, что существуют и существовали специфические ассоциации, брёвна являлись и являются постоянным компонентом пелагиали. Специфическими видами, связанными с плавником, являются морские уточки Lepadidae, обычно дающие почти 100% биомассы пелагического обрастания (Рис. 6.18).
В другие периоды времени пелагическое обрастание выглядело, по-видимому, совсем иначе. Так, в мезозое в его состав входили колоссальных размеров стебельчатые морские лилии (Рис. 6.19). В пелагическое обрастание входят также саргассовый рыба-клоун Histrio histrio, некоторые рыбы-иглы.

Нектон. Только для тропических биомов свойственны голоэпипелагические рыбы, весь жизненный цикл которых от икры до взрослых особей проходит в эпипелагиали. В более высокие широты некоторые из них заходят летом, когда поверхностный слой воды достаточно прогрет.
Большинство видов нектонных кальмаров также ограничено тропиками. Ареалы большинства голоэпипелагических видов циркумтропические, хотя есть и викарирующие индо-тихоокеанские и атлантические виды. Многие виды нектона питаются в мезопелагиали, в частности её обитатели имеют важное значение в питании тунцов, марлинов и меч-рыб и кашалотов.
Голоэпипелагические виды нектона имеются только среди высших, филогенетически молодых семейств костистых рыб (преимущественно окунеобразных, несколько десятков видов) и акул, причём голоэпипелагические акулы по палеонтологическим данным известны с начала неогена, то же справедливо и в отношении нектонных кальмаров, среди которых активно и хорошо плавающих, всего 6 родов с 1–2 видами каждый.
Это свидетельствует о весьма недавнем формировании современного нектона. Н.В. Парин предполагал, что в палеогене была своя голоэпипелагическая биота, но частично она вымерла без следа, и частично была вытеснена в батипелагиаль. При этом какие-либо свидетельства того, что палеогеновая голоэпипелагическая биота вообще существовала, отсутствуют, он просто полагает, что коль скоро был биотоп, должны были быть и специализированные виды.
Не удивительно, что относительно кальмаров К.Н. Несис придерживается противоположного мнения: он считает, что после вымирания белемнитов в конце мезозоя – раннем кайнозое, нектонных видов головоногих не было до сравнительно недавнего времени.В тропических районах имеются специфические формы нектона. Наиболее известны рыбы-лоцманы и рыбы-прилипалы. И те, и другие используют крупных особей нектонных видов почти исключительно для снижения энергетических трат на плавание, а прилипалы — и на дыхание.
Многие виды рыб-прилипал связаны с определёнными таксонами или даже видами. Сопровождают крупные объекты для снижения собственных энергозатрат на плавания не только рыбы-лоцманы, но и такие прекрасные пловцы, как тунцы (известны случаи сопровождения ими китовых акул) и дельфины (широко известны факты сопровождения ими кораблей).
Стабильность биотопа и сложность биома обеспечивают высокую стабильность высшего трофического уровня — нектона. Например, межгодовые колебания численности рекрутов тропических тунцов на порядок меньше, чем у бореальной трески.
В мезопелагиали постоянно обитает разнообразная фауна (Рис. 6.19). Одни виды постоянно находятся в мезопелагиали и питаются мигрирующими видами, другие, как, например алепизавры Alepisaurus и кинжалозубы Anotopteridae поднимаются к поверхности, где питаются эпипелагическим нектоном.


















