Метод выделения конкретных биот
Предложен А.И. Толмачевым и С.М. Разумовским и, на первый взгляд, является вариантом метода типизации ареалов. Однако, различно как применение этих методов, так и интерпретация полученных результатов. Различие заключается в том, что при типизации ареалов работают непосредственно с ареалами видов.
Из-за неполноты данных анализируемые ареалы обычно меньше реальных, причём различия эти неодинаковые у разных видов и в разных участках границ ареала. В результате границу между регионами получают как область сгущения границ ареалов, в пределах которой одна биота плавно сменяет другую.
Действительная граница между биогеографическими выделами представляет собой линию пренебрежимо малой ширины — синперату. Различия между этими подходами иллюстрирует Рис. 2.25.
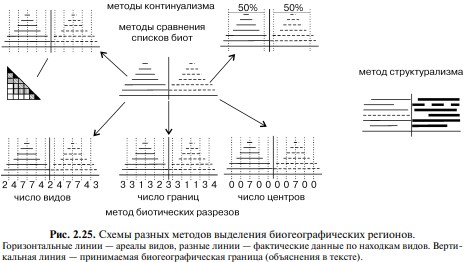
Идея рисунка заимствована у R. Hengeveld. При выделении конкретных биот виды относят к той или иной ЦС (= конкретной биоте), границу которой и считают границей биогеографического региона.
Если в методе типизации ареалов на карту наносят ареалы отдельных видов, то в данном методе на одну карту наносят совокупный ареал видов одной ЦС, имеющих границу ареала в исследуемом районе.
Помимо просто количественного увеличения точек на такой карте из-за суммирования данных по нескольким видам, происходит неизбежное устранение кружева ареала. Причина этого в том, что при достаточном числе видов в это число неизбежно войдут виды разных ассоциаций и сукцессионных рядов, образующих одну ЦС, т. е. виды с разным кружевом ареала, но с общей границей ареала.Как уже говорилось ранее, границу ареалов отдельных видов проводят на основе эктра- и интерполяции, используя для этого карты физико-географических параметров, которые считают значимыми для данного вида (глубина, высота, температура и т. п.). В данном методе для ценофильных видов для этой цели используют ареалы других видов, входящих в ту же ЦС.
Поэтому метод выделения конкретных биот можно также рассматривать как вариант метода типизации ареалов, в котором экстра- и интерполяцию проводят не по одному или немногим произвольно выбранным физико-географическим факторам, а по биотическому окружению вида. Использовать данные по биоте предлагает и К.Н. Несис в приведённой выше характеристике метода типизации ареалов.Принципиально важно, что, поскольку ЦС сменяют одна другую в пространстве, ареалы соседних ЦС комплементарны друг другу. Аналогичного требования нет в методе типизации ареалов. Поскольку границы типов ареалов видов (ЦС) проводят этим методом независимо друг от друга, их комплементарность является независимым подтверждением правильности их проведения, превращая ареал из «произвольной картинки» в верифицируемую гипотезу.
Метод мало чувствителен к слабой изученности распространения отдельных видов и не требует генерализации данных до проведения типизации ареалов.
Использование метода:
- Стадии идентичны соответствующим стадиям метода типизации ареалов.
- После типизации ареалов строят карты каждого типа ареала, на которые наносят полностью находки всех видов с данным типом ареала.
- Границу такого обобщённого ареала и считают границей ареала видов с данным типом распространения.
- Виды, распространение которых сильно отличается от остальных, считают кандидатами в ценофобные, окончательное их отнесение к ценофобным производят после того, как удаётся убедиться в том, что нет видов с ареалами, комплементарным их ареалу.
- Биогеографическими считают только границы комплементарных ареалов. Фактически это означает, что их проводят как общие границы ареалов взаимоисключающих видов. Прочие границы считают лишь границами кружева ареала, но таких границ получается немного, или они вовсе отсутствуют, поскольку большая часть лакун кружева ареала заполняются попутно, ибо они различны у видов разных ассоциаций.
- При правильно проведённом районировании выделы должны покрывать всю рассматриваемую акваторию (территорию).
Поясню различие методов типизации ареалов и выделения конкретных биот на примере анализа наших данных по распространению в Баренцевом море полихет. Пусть нас интересует положение границ ареалов бореальных и арктических шельфовых видов. При получении этих данных были полностью переопределены все коллекционные материалы ЗИН РАН, кафедры гидробиологии МГУ, Зоомузея МГУ и некоторые другие.
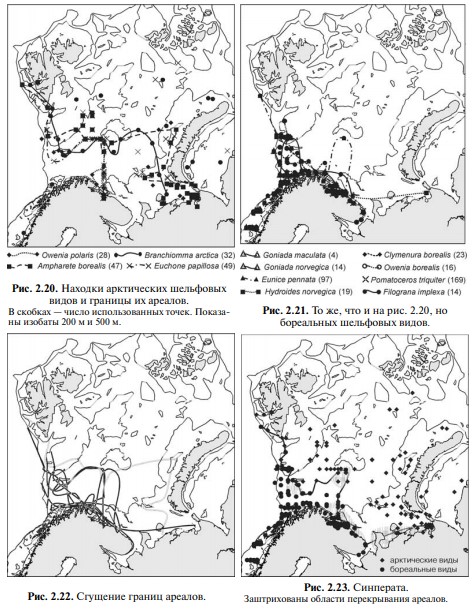
Литературные данные не использованы из-за слишком частых неверных определений, но и без того в анализе использованы сотни точек (см. Рис. 2.20 и Рис. 2.21). На Рис. 2.20. приведены границы ареалов арктических шельфовых видов, проведённые соединением крайних точек находок видов.
На Рис. 2.21 приведены аналогичные границы ареалов бореальных шельфовых видов. На обоих картах границы образуют сложную картину причудливо извивающихся и пересекающихся линий. На рис. 2.22 границы ареалов обеих групп наложены: картина ещё более усложнилась.
Проведём ту же операцию методом выделения конкретных биот. Для этого на карты нанесём все находки арктических и бореальных шельфовых видов. На рис. 2.23 совмещены все данные и показана граница ареалов арктических и бореальных шельфовых видов.
Та же самая граница была нами получена на основе анализа распространения иглокожих по данным А.А. Шорыгина (Рис. 2.24). И здесь получена чёткая граница между арктическими и бореальными шельфовыми видами.

Сравнение карт на Рис. 2.20 – Рис. 2.23 показывает, что область сгущения границ ареалов на самом деле, по-видимому, является областью допустимых положений синператы. Зона вместо линии получается из-за неточностей проведения границ ареалов отдельных видов.
В частности, если метод А.А. Шорыгина (Рис. 2.19) показывает широкую зону перекрывания ареалов арктических и бореальных видов, то метод выделения конкретных биот на тех же данных (Рис. 2.24) показывает, что на больИмеющиеся области перекрывания ареалов, вероятнее всего связаны с тем, что этот метод выявляет в первую очередь границы провинций, т. е. групп ЦС.
Другая причина — зоны перекрывания в открытом море расположены в акваториях со сложным рельефом: возможно здесь на самом деле нет перекрывания ареалов, а лишь сложная мозаика. К сожалению, данных для выявления такой мозаики недостаточно, а у тех, что есть координаты определены с большой погрешностью (по моему опыту в рейсах — до 15`–30` и даже больше), поскольку в то время их определяли счислением, т. е. оценкой положения судна по скорости его хода, известной скорости течений и координатами положения судна, определёнными несколько дней назад в минуты редких просветов в облаках.Важно отметить, что синперата выявляется в результате анализа видов, предпочитающих самые разные грунты (от ила до камней) и обладающих самыми разными типами питания (от фильтраторов до грунтоедов и хищников), т. е. она ни в коем случае не является границей каких-либо групп видов со сходной биологией, а именно границей ЦС.
Дополнительным и неожиданным результатом оказалась необычайная стабильность положения биогеографической границы во времени. На приведённых картах суммированы данные, начиная с конца XIX в. За этот период, как хорошо известно, наблюдались периоды потепления и похолодания.
На самом деле ничего подобного нет. По-видимому, это связано с неправильной интерпретацией данных. Как показано выше, не существует точных методов проведения границ ареалов. В работах, где анализировали распространение видов в Баренцевом море, границы ареалов проводили по крайним точкам нахождения видов. Этот способ, за редким исключением, даёт уменьшенные ареалы видов.
Поскольку вероятность обнаружения вида зависит от его обилия, то при снижении обилия в данном месте, вид уже практически невозможно обнаружить, но это вовсе не означает, что его тут нет. На изменение абиотических условий в пределах экологической амплитуды ЦС (в данном случае — температуры) донные ЦС, имеющие многие черты пастбищных, реагируют изменением относительного обилия их членов.
Поэтому обнаруживаемые «пульсации» ареалов, судя по нашему анализу, вероятно, являются лишь отражением изменения частоты встречаемости этих видов в пределах их ареала, а не самих границ ареалов. Границы между ЦС (т. е. биогеографические границы), как показывают наши данные, весьма устойчивы. По-видимому, для изменения их положения необходимы гораздо более продолжительные климатические изменения, по крайней мере, существенно превосходящие по продолжительности время жизни особей видов-доминантов, которое у видов макробентоса часто составляет многие годы, а у некоторых видов и десятки лет.Незначительные изменения абиоты, по всей вероятности, не столь существенны, поскольку ЦС, а не экологические амплитуды изучаемых видов, обычно определяют границы ареалов видов. Виды же обычно способны нормально развиваться и размножаться в условиях, значительно отличающихся от существующих в их ареале.
Исследование проводят методом последовательных итераций. Данный метод — это первая итерация, поэтому отсеиваемые виды не ценофобы, а только «кандидаты» в них. Хотя, конечно, это может произвести впечатление подгонки результатов путём отсеивания не нравящихся данных.
В какой-то мере от субъективности предохраняет использование двух независимых критериев: нет видов со сходными ареалами и нет видов с комплементарным ареалом. Из теории структурализма следует, что ценофобы обычно удовлетворяют обоим критериям, а ценофилы — нет.
Как дополнительный критерий ценофобности вида можно использовать данные о его распределении. Мозаичность распределения свойственна и ценофильным, и ценофобным видам. Но ценофильные виды обильны в определённых ассоциациях = имеют высокую верность (встречаются обычно совместно в определённых ассоциациях, входящих в ту же ассоциацию), а сама ассоциация приурочена к определённым абиотическим условиям.
В тех случаях, когда разные части биогеографического региона имеют сильно различающиеся абиотические условия, существует вероятность разделения одного биогеографического региона на два.
Проведение границы возможно только в районе, где встречаются специфические для контактирующих ЦС виды.
Легче всего этим методом можно выделить регионы ранга провинции и выше. Для выделения более мелких подразделений, в том числе непосредственно ЦС, необходим анализ распространения большого числа видов, но и в этом случае окончательное решение может быть сделано только после изучения сукцессий.
Примеры использования метода. Метод широко использовал С.М. Разумовский и его последователи. В районировании бентоса он использован нами впервые на разобранном выше примере распространения иглокожих и полихет Баренцева моря, при проведении районирования Северного Ледовитого океана на основе распространения полихет, впрочем, ранняя работа К.Н. Несиса сделана скорее этим методом, нежели методом типизации ареала.
Практически идентичный метод использовал К.В. Беклемишев при районировании пелагиали Мирового океана по распространению зоопланктона: «для биогеографического деления послужили не ареалы непосредственно, а обобщённые карты основных типов ареалов… все ареалы каждой группы были нанесены на одну карту».
Отличие его метода состоит лишь в том, что он проводил не одну общую границу, а три:
- за пределами которой нет ни одного нахождения видов, обладающих ареалом данного типа («0%»);
- по обе стороны от которой проходит равное число границ ареалов («50%»);
- в пределах которой встречаются все виды с данным типом ареала («100%», или «ядро» данной группы видов).
Причиной того, что биогеографические границы в пелагиали приходится определять статистически, является то, что поперёк границы происходит проникновение не только населения соседних регионов (как это идет на суше и в бентали), но и самого биотопа, а также то, что картина распределения течений меняется настолько быстро, что имеющиеся данные не позволяют получить одномоментную картину распределения видов.
Приходится обрабатывать данные за сравнительно длительный период времени, когда перемещение биогеографических границ в пространстве становится заметным.
Правомочность такой замены более чем сомнительна. В литературе часто приводят даже конкретные цифры площадей выявления, а затем их сравнивают полагая, что сравнение списков территорий, равных «площади выявления», даст объективную биогеографическую картину. Подобный подход основан на форме кривой зависимости числа обнаруженных видов от обследованной площади.
Эта зависимость в пределах ареала конкретной биоты должна выходить на плато. Такая форма зависимости действительно следует из того факта, что число видов конкретной биоты имеет предел. При охвате исследованиями ареала соседней конкретной биоты число обнаруженных видов вновь начнет расти.
В дальнейшем кривая будет опять выходить на плато, соответствующее числу видов двух обследуемых конкретных биот. При добавлении данных по ареалу ещё одной конкретной биоты опять будет происходить рост числа обнаруживаемых видов и т. д.Однако для определения формы кривой необходимо, чтобы некоторое число точек в начале кривой было получено по данным, полученным в пределах ареала одной конкретной биоты. Если же уже первые точки взяты в ареале нескольких конкретных флор, то обнаружить площадь выявления элементарной, а, возможно, и нескольких конкретных биот не удастся.
Т. е. разрешающая способность метода зависит от того, насколько представления исследователя о минимальных площадях выявления и, что не менее важно, их форме, соответствуют реально существующей в природе картине.
Более того, метод использует в неявном виде два допущения: распределение видов в пределах биогеографического выдела случайно и что форма биогеографического выдела близка по форме к анализируемому участку.


















